
ホーム > 研究紹介 > 刊行物 > 森林総合研究所関西支所年報第41号 > 年報第41号 主要な研究成果
ここから本文です。
年報第41号 主要な研究成果
1.銀閣寺山国有林の広葉樹二次林における6年間の林相の変化
伊東宏樹(造林研究室)
1. はじめに
放置された広葉樹二次林では遷移の進行により、常緑広葉樹が増加しつつあるといわれる。しかしながら、こうした林分は近年になるまで生態学的な調査の対象としてあつかわれることは少なく、その実態を継続調査した例も多くない。本報告では、京都市内の放置二次林を対象として6年間にどのような変化が起きたかについて報告する。
2. 調査地及び調査方法
調査は、京都大阪森林管理事務所管内の銀閣寺山国有林(京都市左京区)に設置した固定調査区で行われた。1992年に1.05haの面積の調査区を設置し、1993年・1996年・1999年に胸高直径3cm以上の木本及び藤本について毎木調査を実施した。調査区の標高は140~210m、母岩は花崗岩であり、南向き斜面である。調査区内には一部にヒノキ人工林を含んでいたが、この部分は1999年に土砂崩れを起こしたため調査が不能となった。このため、天然林部分の5000m2について解析した。
3. 結果及び考察
天然林部分の5000m2の調査区について、1993年には46種が確認され、幹密度は3314本/ha、胸高断面積は35.4m2/haであった。胸高断面積比でみてもっとも優占していたのはクロバイ(Symplocos prunifolia Sieb. et Zucc.)で、以下タカノツメ(Evodiopanax innovans (Sieb. et Zucc.) Nakai)、アラカシ(Quercus glauca Thunberg)、アオハダ(Ilex macropoda Miq.)、コナラ(Quercus serrata Murray)の順だった。
1999年には、種数・幹密度・胸高断面積はそれぞれ、50種、3088本/ha、39.1m2/haとなった。6年間で、幹密度はおよそ6.8%減少し、胸高断面積は10.5%増加したことになる。
この間の枯死木を樹種別にみると、タカノツメがもっとも多く、1993年に生存していた308本中89本が1999年までに枯死していた。ついでアラカシ29本、フジ(Wisteria floribunda (Willd.) DC.)20本、クロバイ18本の順で、タカノツメの枯死がきわだって多かった。一方、この間に新規加入した樹種としては、サカキ(Cleyera japonica Thunberg)が37本で最も多く、ついでアラカシ33本、ヒサカキ(Eurya japonica Thunberg)12本、クロバイ10本の順だった。
6年間の、落葉広葉樹と常緑広葉樹のそれぞれの胸高直径階別の幹数の変化を図-1に示す。落葉広葉樹の小径木の減少が顕著である。一方、常緑広葉樹は安定した齢分布をたもっている。このまま放置が続き、大規模な撹乱がなければ、落葉広葉樹の小径木はますます減少し、高木層にある落葉広葉樹が枯死した後は、アラカシなどの常緑広葉樹が中心の林分になるものと予想される。ただし、本調査区では、先に述べたような規模な土砂崩壊がしばしば起こるため、こうした撹乱がどのように遷移に影響を与えているかについても考慮する必要がある。
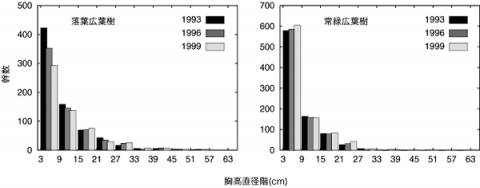
図-1 落葉広葉樹と常緑広葉樹の胸高直径階別の幹数の変化
2.アラカシ実生の植栽試験
伊東宏樹(造林研究室)
1. はじめに
アラカシは、乾燥や撹乱に強いなど常緑広葉樹としては先駆種的な性格を持つとされる。実際、放置された広葉樹二次林でアラカシが増加しているという報告が各地でなされている。銀閣寺山国有林の広葉樹二次林に設置した固定調査区でも、多くのアラカシが見られる。
この調査区では、アラカシの成木は、急斜面や斜面中部?下部を中心として密度が高く、一方で尾根部には低いという、地形と対応する分布パターンを形成している。このパターンは、林床の稚樹・実生についてもほぼ同様である。こうした分布パターンが形成された要因として考えられるもののうち、特に実生の定着初期に関わるものとして、(1)母樹の分布パターンを反映した種子散布量の違い、(2)地形による発芽率の違い、(3)地形による実生定着率の違い、の3つが考えられた。そこで今回は、このうちの(3)の可能性を検討するため、植栽試験を実施した。
2. 試験区および試験方法
銀閣寺山国有林(京都大阪森林管理事務所管内、京都市左京区)の広葉樹二次林と、京都府南部の京田辺市の広葉樹二次林に1m2の面積の試験区を、銀閣寺山では尾根・斜面上部・斜面下部・谷の4カ所、京田辺では尾根・谷の2カ所の合計6カ所に設置した。銀閣寺山試験区の母材は花崗岩、京田辺試験区の母材は第四紀更新統堆積物である。すべての試験区で林冠はほぼ閉鎖していた。林冠木の樹種は、銀閣寺山がアラカシ・タカノツメ・クロバイ・ソヨゴなど、京田辺がアカマツ・コナラ・ソヨゴなどであった。
1998年春に、京田辺市内でアラカシの種子を採取し、これを発芽させ、ポットで育苗した。これらのうちから、発育状況が著しく異なる個体をのぞいた96個体を選び、1998年7月上旬に1試験区あたり16個体のアラカシの実生を植栽した。その後、継続的に状況を観察し、1999年秋までの生存個体数に処理間で差があるか検討した。
3. 結果および考察
1999年秋までの生存数を表-1に示す。銀閣寺山の谷試験区は、1998年8月にイノシシが地面を掘り起こしたことより、根が掘り出されたり、植物体が土に埋もれたりするなどして、同年秋までに13個体の実生が枯死した。このためこの処理区のデータは解析に用いなかった。
その他の試験区について、平均の生存率(0.6125)によって期待される生存数と、観察された生存数との間に差がないとの帰無仮説の元で、2項検定により検定を行った。その結果、すべての試験区で、試験区間での有意な差は認められなかった。
この結果から、少なくとも今回の試験地のような場所では、アラカシの実生の生存に地形要因は大きな影響を及ぼしているとはいえない。したがって、銀閣寺山国有林の固定調査区にみられるアラカシ稚樹・実生の地形と対応した分布パターンの形成要因としては、はじめにあげた3つのうち(1)または(2)によるものである可能性が高いものと考えられる。
| 場所 | 地形 | 植栽数 | 生存数 | 備考 |
| 銀閣寺山 | 尾根 | 16 | 9 | |
| 斜面上部 | 16 | 7 | ||
| 斜面下部 | 16 | 13 | ||
| 谷 | 16 | — | イノシシの掘り起こしによる被害 | |
| 京田辺 | 尾根 | 16 | 11 | |
| 谷 | 16 | 9 |
3.シカとササの除去が土壌の養分状態に及ぼす影響
古澤仁美(土壌研究室)
1. はじめに
奈良県にある大台ケ原の林床ではミヤコザサ(以下、ササという)が優占しており、ササと樹木実生とは光だけでなく土壌中の養分をめぐる競争関係にあると考えられる。しかし林床のササが土壌中の養分状態へ与える影響は明らかになっていない。一方、大台ケ原にはササを主食とするニホンジカ(以下、シカという)が高密度で生息している。シカはササを採食することで土壌の養分状態に変化をもたらし実生へ間接的な影響を及ぼしていることも考えられる。そこで森林内でシカ、ササを実験的に制御した試験地において、水溶性イオンを土壌の養分状態の指標として測定し、水溶性イオンへ与えるシカ、ササの影響を検討した。
2. 研究方法
大台ヶ原山のブナ、ウラジロモミを主要構成樹種とする針広混交林にシカ、ササの2要因の除去操作の有無を組み合わせた4とおりの処理区からなる実験区を設置した。この4処理区から1997年~1999年の春(5月ないし6月)、及び秋(9月ないし10月)に無機土壌(0-5cm)を採取した。土壌は採取後直ちに低温で研究室まで運送し、冷蔵保管したのち2mmメッシュのふるいを通して供試土壌とした。供試土壌1に対し水5を加えて振とうし、ろ過した抽出液の水溶性イオン濃度をイオンクロマト法で分析した。各々のイオンについて各々の測定時期ごと、及び3年分の秋のデータについて分散分析法でシカ、ササ処理の影響を解析した。
3. 結果と考察
すべてのイオンについて処理の影響は春には認められず、秋に認められた。そこで、3年分の秋のデータを用いて分散分析をした結果を表-1に示した。アニオンのうちNO2-でシカ、ササの負の効果が認められた。NO3-ではササの負の効果が認められた(図-1)。カチオンではNH4+でシカの負の効果が認められ、Ca2+、Mg2+ではシカの正の効果及びササの負の効果が認められた。ササの負の効果はササがこれらのイオンを吸収しているために生じると考えられ、林床のササと実生が養分をめぐって競争関係にあることが確かめられた。処理の影響が秋のみ認められるのは、春は生物が活動を開始してまもないため差があらわれないが、秋までに生物活動の効果が蓄積されるためと考えられた。シカの正の効果があらわれるのは、シカがいる方がササの現存量が小さいためと考えられ、シカも土壌中の水溶性イオン濃度に影響を与えうることが示唆された。しかし、NH4+でシカの負の効果がなぜあらわれるのかは不明であり今後検討していく必要がある。
| シカ | ササ | シカ×ササ | |
| Cl- | ns | ns | ns |
| (p=0.397) | (p=0.811) | (p=0.608) | |
| NO2- | - | - | ns |
| (p=0.000) | (p=0.000) | (p=0.108) | |
| NO3- | ns | - | ns |
| (p=0.611) | (p=0.000) | (p=0.385) | |
| SO42- | ns | ns | ns |
| (p=0.076) | (p=0.342) | (p=0.228) | |
| Na+ | ns | ns | ns |
| (p=0.581) | (p=0.158) | (p=0.550) | |
| NH4+ | - | ns | ns |
| (p=0.000) | (p=0.196) | (p=0.242) | |
| K+ | ns | ns | ns |
| (p=0.817) | (p=0.084) | (p=0.555) | |
| Ca2+ | + | - | ns |
| (p=0.000) | (p=0.000) | (p=0.990) | |
| Mg2+ | + | - | ns |
| (p=0.000) | (p=0.000) | (p=0.868) |
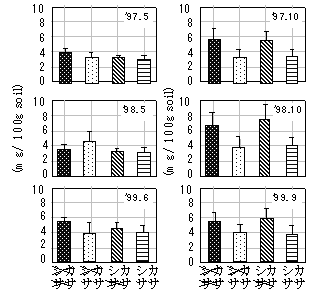
図-1 NO3-濃度に及ぼすシカ、ササ処理の影響
4.堆積有機物層からの溶存有機炭素発生量のシミュレーション
南部桂(科学技術特別研究員)
金子真司(土壌研究室)
1. はじめに
溶存有機物は土壌表層の物質移動や炭素サイクルに大きな役割を果たすものとして注目されているが、その発生量がどのように決定されるかについては十分に理解されていない。室内実験からは、「高温であるほど、また抽出回数が多いほど発生量が多い。培養初期に発生量が多い」という結果が得られているが、野外においてははっきりした季節変化はみられず、実験結果との間の整合性は確かめられていない。そこで、本研究は培養実験で得られる結果に基づいて溶存有機炭素(DOC) 発生量の予測式を作り野外の観測値と比較することにした。
2. 方法
DOCは植物遺体が分解されるときに生じる分解の中間産物であるので、有機物の分解量の経時的変化に関して用いられるモデル式(1次反応式やロジスティック式、及びそれらの組み合わせた式)を当てはめることにした。また、温度が与える影響についてはアレニウス則を用いて解析した。培養実験A: 落葉直後の京都市北部ミズナラ林のF層を採取し、30, 25, 20℃の3水準で培養しながら10日おきに100mlの脱塩水で抽出してDOCを採取した。培養実験B: 上記のF層を30℃で培養しながら、3.5, 10, 28日おきの3水準の間隔で100mlの脱塩水を用いて抽出した。実験C: 翌年の4月に再度野外からF層を採取し、実験Aを繰り返した。この実験で得られたDOC発生量は、作成された予測式の当てはまりを確認するための比較用実測値として用いた。野外観測: A0層直下の浸出液をテンシオンフリーライシメータで採取した。浸出液のDOC濃度から林内雨のDOC濃度を差し引いた後、林内雨量を掛け合わせて、DOCの野外における発生量とした。また、野外の日雨量および日平均気温をモデル式のインプットデータとした。
3. 結果と考察
培養温度が高いほど(実験A)、また抽出間隔が長いほど(実験B)各回の抽出におけるDOC発生量は多かった。しかし、この傾向は培養初期(約40日目まで)に顕著であったが、それ以降は差が縮まり100日目以降は処理によらず有意な差は見られなかった。これは、DOCが分解の産物であるため微生物活性に影響されることと、培養の初期にはDOCの発生源となる基質が豊富であったが日数とともに減少したことを示し、一般的な有機物分解モデルが当てはまるものと思われた。一方、実験Bで同じ培養期間におけるDOC発生量の積算値を比較したところ、抽出回数の多い処理ほど積算発生量は多かった。これは、分解反応とは無関係に抽出操作の回数がDOC発生量に影響を与えたことを示す。実験Aの結果から、「分解に伴う過程」と「抽出操作に由来する過程」から成るモデル式を作成し、パラメータを推定した。前者の過程は2つの1次反応式の2段階並行式で、後者の過程は抽出回数に比例する式で表された。この式で実験B及びCの結果を予測したところ、十分な精度で予測できた。推定式を用いて予測した野外の発生量と実測値を図-1に示す。予測値(太い実線)は5/6まではほぼ正確であったが、その後は約2/3の過小評価にとどまった。過小評価になった原因はL層からのDOC発生を考慮しなかったからであろう。仮にL層からの発生量もF層と同様であると仮定すると、予測値は点線となり4月以降の実測値とよく合致した。L層は積雪によって圧密化を受けることによってより湿潤な状態が保たれやすくなることが現場で観察された。このことによって融雪期以降はL層もDOC発生により寄与するようになったのではないかと考えられた。
「分解過程」によるDOC発生量は4月~7月に最も多かったがそれ以降は基質が少なくなって減少した。落葉後~積雪期は低温のため最も少ないと予測された。一方、「抽出過程」による発生量は本地点では年間を通して大きな変化はなく、また全発生量の6割程度を占めるものと計算された。
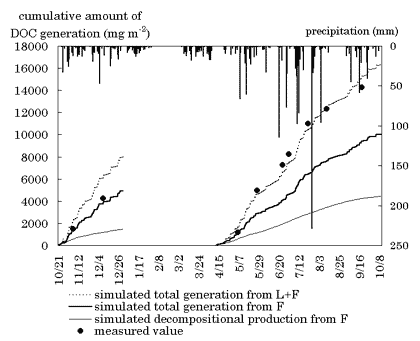
図-1 DOC発生量の計算値と実測値の比較
5.京都市山科区の森林流域における物質収支の特徴
金子真司(土壌研究室)
1. はじめに
関西支所土壌研究室では、特定研究「酸性雨等の森林生態系への影響モニタリング」の一環として、京都市内のスギ林において降雨、林内雨、樹幹流及び渓流水の水質調査を行っている。ここでは調査流域における物質収支の特徴を明らかにすることを目的として、降雨から流域への物質負荷量と流域から渓流水への物質流出量を調べ、京都市と同緯度に位置する米国の森林流域の物質収支との比較を行った。
2. 調査地及び方法
京都市山科区安祥寺山国有林における1995年から1999年の5年間の降雨と渓流水質データに基づき、物質の流入と流出量を計算した。本調査地では流量観測は行っていないので、蒸散量は京大桐生試験地の報告を参考に828.8mm/yearであると仮定し、各年の降水量から828.8mmを引いた値を年流出量とした。比較には米国ノースカロライナ州カウィータ水文試験地のWS2流域におけるデータを用いた。ここでは前者を山科、後者をカウィータと呼ぶことにする。山科における渓流水の採取地は北緯度35°00′13549、東経135°49′標高150mで、流域の最高地点は460m、採取地から上流の流域面積は約100haである。流域の大半はスギ・ヒノキの人工林で覆われており、地質は中・古生層堆積岩である。カウィータは北緯35°03′、西経83°25′に位置し、全流域の標高は675~1592m、年平均降水量は1771mmで流域の大半は落葉広葉樹林に覆われている。地質は先カンブリア時代の堆積岩である。
3. 結果及び考察
図-1に山科とカウィータの降雨からの成分負荷量(インプット)を示した。山科はNa+とCl-の負荷量がカウィータの約4倍、またMg2+の負荷量が約2倍であったが、他の成分の差は小さかった。山科はアメリカ大陸内部のカウィータに比べて海に近いために、Na+、Cl-、Mg2+の負荷量が大きかったと考えられる。成分流出量(アウトプット)に関しては、いずれの成分も山科の方が大きかった(図-2)。特にMg2+、Ca2+、Na+のアウトプットはインプットの7.6、6.1、3.9倍であったことから、山科では岩石の風化によって供給される物質量が多いと推定される。酸性雨の原因物質と言われているSO42-とNO3-は、カウィータではアウトプットがインプットに比べて極端に小さかったのに対して、山科ではアウトプットがインプットを上まわっていた。NO3-は植物の必須元素であり一般の森林では窒素不足であるためにNO3-の流出は少ないと言われている。SO42-とNO3-は一般に岩石から供給されないので、アウトプットがインプットより多くなることは考えにくい。少なくとも山科では降雨以外のNO3-とSO42-の供給源(例えば樹木への乾性沈着)があると推察される。
山科とカウィータはほぼ同緯度にあり、気候条件も類似している。流域は傾斜があり、森林に覆われており、氷河による剥離も受けていない。このように良く似た条件の流域であるが、物質収支、特にアウトプット量に著しい違いが認められた。カウィータではアルティソル(赤黄色土)が分布していることから、キャンビソル(褐色森林土)が分布する山科に比べて土壌生成期間が長いと推定される。そのためにカウィータでは溶存物質量が少なくなっている可能性が考えられる。NO3-とSO42-のアウトプットの違いについては今後さらに詳しい検討が必要であろう。
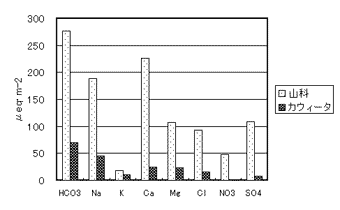
図-1 流域間の負荷量(インプット)の比較
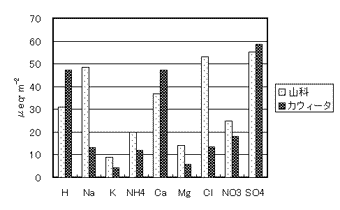
図-2 流域間の流出量(アウトプット)の比較
参考文献:Swank TS & Crossley DA (1988) Forest Hydrology and Ecology at Coweeta, Springer-Verlag.
6.木造3階建て住宅における木材の使用とその供給
―長尺の通し柱を中心に―
野田英志(経営研究室)
1. はじめに
木造3階建て住宅は、昭和62年の建築基準法改正で準防火地域内での建築への道が開かれて以降、大都市部を中心に建築が増大してきた(平成元年度0.6万棟→10年度3.3万棟)。とりわけ京都・大阪の大都市部においては、狭い敷地で住宅建築が可能になることなどの理由から建築が増加し、平成10年度には1.3万棟と、地区内の木造住宅建築戸数の3割前後を占めるまでになっている。工法別には軸組が59%(10年度・全国)と過半を占める。とくに京都府では軸組の割合が97%と高い。こうした木造軸組の3階建て住宅には一般に、1階から3階に至る長尺の通し柱(長さ8~9m)が使用される。木造軸組3階建て住宅に特有の部材であるこの長尺の通し柱は、木材新商品ともいえるが、その使用実態や供給仕組みについては不明な点が多い。そこで本研究では、近年、関西地域で増加が著しい木造3階建て住宅に注目し、そこで使用される長尺の通し柱にスポットを当てて、その使用実態と供給の仕組みを探った。
2. 京阪地区の木造3階建て住宅における木材使用事例 ―長尺の柱材を中心に―
京阪地区都市部の木造3階建て住宅(主に建売)の特徴は、概ね洋風調のカラーベスト総3階建て・敷地50~70m2・延べ床面積100m2前後(建ぺい率60%、容積率200%)で、道路に面して間口の狭い短冊状の狭小敷地いっぱいに建て、1階にガレージのスペースをもつ住宅が多い。平成10~11年に調査した建築中の木造3階建て建売住宅団地(京都市及び周辺市部の3ヶ所・計30棟・区画数59)のケースでは、いずれの住宅も3階までの長尺通し柱(長さ9m×120ミリ角)を6本使用している。団地の2カ所がヒノキ芯持で背割のある柱、1カ所が集成柱であった。なお年間50~60棟の木造3階建て住宅部材のプレカット加工を行っている京都市近郊のKプレカット工場の場合、長尺の通し柱は1棟当たり4~6本の使用で、その樹材種の割合はヒノキ7割、集成材3割である。長尺の通し柱以外の管柱や横架材などの構造材は、2階建て木造住宅に使用される材と同じものが使われている。
このように木造軸組3階建て住宅には、4本ないし6本程の長尺通し柱が使用されていることがわかる。ただし道路事情などで長尺材の搬入が困難な現場では、1~2階と2~3階に6m通し柱を使用するケースもある。長尺通し柱の市場規模を上記の結果から推測すると、京阪地区の場合、木造軸組3階建て住宅建築棟数(平成10年度7,478棟)から判断して、年間およそ2~3万本程の市場規模ではないかと見られる。
3. 長尺通し柱材の供給の仕組みと課題
3階建て用の長尺通し柱は、6m通し柱のように既製品として製品市場で流通していない。工務店・地域ビルダーから注文を受けた木材販売店・プレカット工場は、長尺材の加工設備を備えた製材工場や集成材工場へ発注する。
長尺通し柱を生産している大手製材工場(岡山県)の場合、6m材用の台車では長尺材の両端が固定できず挽き曲がりが出るため、長さ10mの材まで製材可能な台車を導入している。同工場では月間2~3百本のヒノキ9m通し柱(KDモルダー加工)を生産。販売先は京阪神地区の木材問屋が6割、同・プレカット工場4割。一般に問屋は在庫は持たずに必要本数のみの注文であり、一方、プレカット工場は数バンドル(12本/バンドル)のロット買いが特徴である。また、9mの長尺素材は原木市場へ注文を出し、市場が出荷者(素材生産業者・森林組合)へ造材を依頼する特注生産の形態をとっている。京都府京北町の原木市場調査によると、出荷者が市場からの依頼なしに自らの判断で9m材を造材して市場に出荷することはなく、製材工場からの依頼を受けた市場がその都度、出荷者に必要本数の造材・出荷を要請している。
ヒノキ9m長尺材は、70~80年以上のヒノキ高齢級人工林から生産される。長尺通し柱の消費地は、京阪地区などの特定大都市地域に集中している点を考えると、高齢級人工林資源を擁する近畿地方の先発林業地(京北町など)は、資源面でもまた立地面においても有利な条件下にあるといえよう。ただし近年、木造3階建て住宅の欠陥・違法建築が社会問題化しており、林業・木材供給サイドの対応が迫られている。こうした状況に対応して、集成材と金物接合を多用した木造3階建て新工法(SE工法など)が開発され、近年、その住宅供給が拡大している。こうした新工法住宅には、品質のバラツキの大きい国産無垢材は採用されない。
近年のこうした新たな状況を踏まえると、先発林業地では旧来の木材供給システムだけに依拠するのではなく、地区内の高齢級資源の高度活用と国産無垢材の市場確保を目指して、1)川下の木造3階建て住宅の建築情報と川上の高齢級林分情報をリンクした択伐方式での長尺材生産システムや、2)高品質の長尺通し柱の製材・乾燥・機械プレカット加工の一貫生産システム、そして3)木造3階建て住宅躯体の構造・強度チェック体制整備などにより、高品質・性能保証された木造3階建て住宅部材の新たな供給システム構築が必要なのではなかろうか。
7.ヒノキ漏脂病被害林分における低温履歴時間の空間変動
玉井幸治(防災研究室)
池田武文(樹病研究室)
1. はじめに
ヒノキ漏脂病の発生原因には、病原菌、昆虫の他に低温も関与しているという指摘もある。そこで樹体温度が、ある基準温度以下であった時間を低温履歴時間と定義し、それを外的基準とする数量化I類によって解析することにより、低温履歴時間の空間変動を明らかにした。そしてヒノキ漏脂病患部の空間分布傾向との関係を検討した。
2. 観測林分と解析方法
石川県金沢市内にある約30年生ヒノキ漏脂病の被害林分において、斜面位置(上・中・下)、樹幹上の位置(0.5, 1.0, 2.0, 3.0, 4.0, 6.0m)、病患部(有・無)を変えた36ヶ所で、1995年12月~1996年5月に樹体温度の測定を行った。低温履歴時間を算定する基準温度には調査全期間を対象に-2.5℃と-1.0℃の2つの温度範囲を設定した。これらよりも温度が低かった時間を外的因子、斜面位置、樹幹上の位置、病患部をアイテムとする数量化I類の解析を行った。
3. 結果と考察
数量化I類による結果を図-1に示す。樹体温度が-2.5℃よりもさらに低下する多くの場合は、数日間にわたって低温状態が継続しており、これは寒波の来襲に起因するものと考えられた。これに対し樹体温度が-1.6~-0.9℃より低下するが-2.9~-2.3℃までは低下しない場合は、主に放射冷却などによると考えられる一時的なものであった。樹体温度が-2.9~-2.3℃よりもさらに低下する場合、斜面上の中・下部と樹幹上の高さ1.0~3.0mの空間で、低温履歴時間が長い傾向にあった。これは、解析を行った2つの基準温度の中ではヒノキ漏脂病患部の分布傾向と最も良く一致した。これは比較的厳しい低温状態がヒノキ漏脂病患部の形成要因の一つである可能性を示している。
参考文献 玉井ら(2000) 漏脂病被害林分におけるヒノキ樹体の低温履歴時間の空間変動特性、日林誌82、200-203.
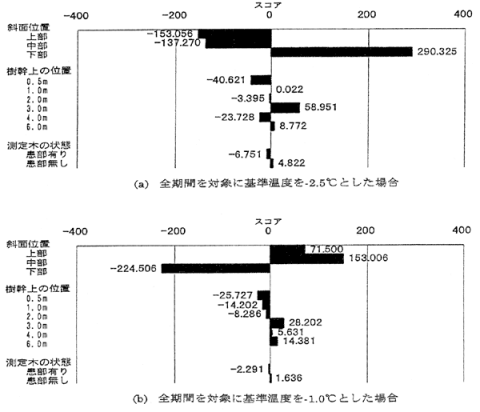
図-1 数量化I類による各カテゴリーのスコア(図中の数値は各カテゴリーのスコアを示す。)
8.緑化を目的としたワラビの植裁条件について
深山貴文(防災研究室)
1. はじめに
ワラビは山菜としての需要がある一方、根茎による無性繁殖が比較的容易なため、在来植物の一種として植生土嚢等への応用も考えられる。このようにワラビを緑化資材として用いる場合、植え付ける根茎長が短いほど根茎の取り扱いは容易であると考えられる。しかし、山菜採集を目的とするこれまでの多くの研究においては食味の改善や促成栽培に主眼がおかれ、植え付けに最低限必要な根茎長等といった厳しい環境での植裁条件は明らかになっていない。そこで、本研究では根茎長の異なる根茎を植え付け、地被がある場合と無い場合のそれぞれについて、無性繁殖に必要な根茎長を明らかにすることを目的とした。
2. 試験地及び試験方法
試験地は京都市伏見区にある森林総合研究所関西支所の苗畑である。苗畑には1m2の試験区を8区設けた。そこへ、京都市山科区の自生地から3時間前に掘り出した直径6mmのワラビの根茎を、5cm、10cm、20cm、50cmと4通りの長さに切断して植え付け、それぞれをマルチあり区とマルチ無し区の2通りで栽培した。1区あたりに植え付けた根茎の総延長は2m、生重量で400gとし、切断した根茎には休眠芽を持つ節が含まれるようにした。マルチには植物繊維マットを用いた。植え付けは1997年4月8日に行い、以後1ヶ月おきに葉長を測定し、新たに発生した葉柄には個体識別用のナンバーテープを付けた。一方、京都市山科区の自生地にも4m2の試験プロットを設け、苗畑と同様の試験を行った。栽培は降雨以外、無冠水で行なった。試験は全てのワラビ葉柄が枯死した1998年1月に終了した。なお、試験終了時にはマルチ無し区の根茎を掘り上げ、1区あたりの生重量と根茎長の合計を求めた。
3. 結果及び考察
根茎長10cm以上の区で葉柄の発生が認められ、植え付けた根茎の長さが長いほど葉柄の発生数が多くなる傾向が認められた。また、各区の平均葉長は10月から12月頃にかけて最大となった。10月の平均葉長を大きい方から並べると50cm+マルチ区>20cm+マルチ区>10cm+マルチ区>50cm区>20cm区>10cm区の順であった。表-1にはt-検定による10月の葉長の検定結果を示した。検定の結果、同じ長さの根茎でもマルチが有ることによって有意に葉長が大きくなることが分かった。図-1には試験終了時におけるマルチ無し区の根茎の生重量と長さの増減を示した。5cm区では全ての根茎が腐朽によって消滅していた。また、10cm区も腐朽した根茎が多く、生重量は植え付け時より減少したが、活着した個体が細い根茎を伸ばしたため、根茎長は植え付け時より増加していた。20cm区と50cm区は生重量、根茎長共に増加しており、休眠芽も多く形成されていた。これらの結果から、十分な活着や良好な成長を期待するには植え付け時に少なくとも20cm以上の根茎長が必要で、地被が存在する事が望ましいと考えられた。
| 順位 | 1 | 2 | 3 | 4 | 5 | 6 | 7 | |
| 1 | 自生地 | |||||||
| 2 | 50cm+マルチ | ** | ||||||
| 3 | 20cm+マルチ | ** | ||||||
| 4 | 10cm+マルチ | ** | ** | * | ||||
| 5 | 50cm | ** | ** | ** | ||||
| 6 | 20cm | ** | ** | ** | * | |||
| 7 | 10cm | ** | ** | ** | * |
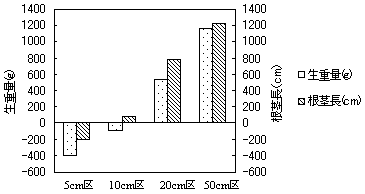
図-1 生重量と根茎長の増減
9.ナラ類集団枯損に関連する糸状菌の成長に対する温度の影響
高畑義啓(樹病研究室)
1. はじめに
ナラ類集団枯損の被害木やカシノナガキクイムシからの微生物の分離結果、及び複数の接種試験の結果から、ある未同定の糸状菌(ここでは「ナラ菌」と仮称する)がナラ類樹木に対して病原性を持ち、集団枯損を引き起こしているのではないかと推測されている。しかしナラ菌は未同定であるばかりでなく、生理学的性質も詳しい報告がない。そこでナラ菌の生理学的性質解明の一端として、培養温度がナラ菌の成長に与える影響について検討を行った。
2. 材料と方法
実験には産地の異なるナラ菌9菌株(表-1)を用いた。PDA平板培地上で培養したコロニーの周縁部を内径4mmのコルクボーラーで打ち抜き、これを直径9cmのシャーレ内のPDA平板の中央部に置き、暗黒下5、10、15、20、25、30、35℃の各条件下で培養した。成長したコロニーの直径を1日ごとに計測し、培養時間に対して直線的な成長が見られる期間における1日あたりの菌糸伸長量を算出した。各菌株・培養温度ごとの反復数は5としたが、コロニーがシャーレ外壁に到達し始めた時期以降では反復数が5未満となった実験区がある。
| 番号 | 採集地 | 分離部位 |
| NA9 | 福井県今庄町 | 枯死木の樹皮 |
| NA21 | 新潟県小千谷市 | 被穿孔木の辺材変色部 |
| NA24 | 兵庫県香住町 | カシナガ雌成虫体表 |
| NA29 | 滋賀県西浅井町 | 萎凋木の孔道壁 |
| NA54 | 京都府福知山市 | 枯死木の辺材変色部 |
| NA56 | 山形県朝日村 | 枯死木の辺材変色部 |
| KA007 | 宮崎県綾町 | アカガシ木部 |
| KA094 | 鹿児島県鹿屋市 | アカガシ木部 |
| KA122 | 鹿児島県内之浦町 | マテバシイ木部 |
3. 結果及び考察
供試した殆どの菌株で、培養温度30℃までは菌糸成長速度が温度の上昇に伴って直線的に増加し、多くの菌株で培養温度30℃での菌糸成長速度が最大となった(図-1)。25℃と30℃での成長の速さの差が統計的に有意でない菌株もあり、菌糸成長速度を指標にした場合のナラ菌の成長適温は25~30℃であると推察された。コロニーの乾燥重量を指標とした場合の成長適温は20~25℃と推察された(高畑、未発表)。一般に、コロニーの拡大の成長適温はコロニー乾燥重量の成長適温より高温側にずれるとされる。このことによって両指標の違いによるナラ菌の成長適温の差は説明可能であるが、より詳細な検討が必要である。培養温度5℃での菌糸成長速度は1.0~4.1mm/dayと非常に遅かったが、成長が停止した菌株はなかった。培養温度35℃では供試9菌株のうち7菌株で成長が停止し、成長が見られた菌株でも成長の速さは1mm/day未満であった。このことから、ナラ菌は35℃を超える温度では生育しないものと推察された。
同一温度における成長の速さには菌株の違いによって統計的に有意な差が認められる場合が数多くあり、菌株の組み合わせによっては成長の速さに2倍程度の差が生じているものもあった(図-1)。このことから、ナラ菌の菌株によって生理的性質が異なる可能性が考えられる。供試した菌株は東北・北陸、近畿北部、鹿児島、宮崎の各地域産に分けられる(表-1、なお、NA9菌株は産地である今庄町のナラ枯損被害地が滋賀県の被害地と隣接しているため、近畿北部産の菌株として取り扱った)。近畿北部産の菌株(NA9、NA24、NA29、NA54)及び宮崎県産の菌株(KA007)、鹿児島県産の1菌株(KA094)は全体に成長が遅く、成長適温である25、30℃においても成長の速さは30mm/day未満であった。また、適温での成長の速さは20℃での値と比べて大きな差がなかった。これに対して、東北・北陸産の菌株(NA21、NA56)と鹿児島県産の1菌株(KA122)は全体に成長が速く、培養温度が20℃を超えても、培養温度の増加に対して直線的な成長の速さの増加が見られた。このことから、鹿児島県産の菌株についてはさらに検討を加える必要があるが、培養温度が菌糸の成長に与える影響について、東北・北陸産、近畿北部産、宮崎県産の菌株の間には産地による差が存在する可能性があると推察された。
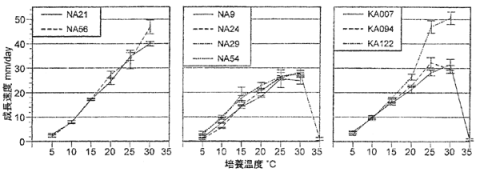
図-1 各培養温度におけるナラ菌9菌株の成長速度
左: 東北・北陸産菌株、中央: 近畿北部産菌株、右: 九州産菌株
35℃にプロットのない菌株はこの温度で成長が停止したことを示す。
10.枯れたマツ由来の各種誘引剤を用いた甲虫類の捕獲調査
上田明良・藤田和幸(昆虫研究室)
1. はじめに
害虫とその天敵の誘引物質を解明することは、害虫の捕獲法や天敵の誘致法開発の指針となる。枯死したマツ類からの揮発成分であるピネン類やエタノールに様々な甲虫類が誘引されることが知られている(上田・藤田 1998「森林応用研究7:113-116」など)。そのうち、マツノマダラカミキリはα-ピネンに誘引されるが、これにエタノールを共用すると誘引力が大幅に増強することが知られている(池田ら 1986「日林誌68:15-19」など)。これに対し、クロカミキリではエタノールがピネン類の協力剤として作用しないことが判明している(Ikeda et al. 1979「Appl. Ent. Zool. 15: 358-361」)が、その他の甲虫類についてはエタノールの作用が調べられていない。そこで、様々な甲虫類についてエタノールの誘引剤としての作用を調べた。
2. 研究方法
1999年4月27日~6月8日の間、滋賀県野洲町近江富士花緑公園内の2~4m高のアカマツの枝にサンケイ式昆虫誘引器(黒色)を改良した関西式生捕り用誘引トラップ(細田ら 1995「日林関西支論4:171-172」)を設置した。誘引剤として、α-ピネン及び(-)β-ピネン単体とそれらのエタノールとの共用、エタノール単体、および対照として蒸留水の計6種を用い、6~8日間隔で誘引剤の交換と捕獲虫の回収を行った。
3. 結果と考察
全部で55種2879頭の甲虫が捕獲され、このうちの14種では回収日ごとの各誘引剤への捕獲数から算出した捕獲順位に有意差が見られた。今回の結果とマツノマダラカミキリ(池田ら 1986)およびクロカミキリ(Ikeda et al. 1979)の食性と誘引物質の関係を表-1に示した。害虫としてあげられるマツ内樹皮食性のものの多くはピネン類に誘引され、エタノールが協力剤として作用することが分かる。ニセマツノシラホシゾウムシは(-)β-ピネン、フィリピンキクイムシとマツノツノキクイムシはα-ピネンとの共用で多かった。捕食者のうちマツ類に特化したオオコクヌストとカッコウムシ2種はα-ピネンと(-)β-ピネンの両方が誘引物質で、エタノールが協力剤として作用していた。これに対し、マツ類に特化していない捕食者の一部や菌食者の一部ではエタノールが誘引物質で、これらはピネン類に影響されないものとピネン類によってエタノールの誘引効果が下げられるものとがあった。ピネン類とエタノールの関係をより明確にするためには、今後様々な場所で長期間の捕獲を行い、比較できる種数を増やすことが必要である。
| 表-1 食性と誘引物質の関係 | ||||
| 誘引物質 | 共用剤の影響 | マツ内樹皮食性 | 捕食性 | 食菌性(含養菌キクイムシ) |
| ピネン類 | エタノールが協力 | マツノマダラカミキリa | オオコクヌスト | |
| ニセマツノシラホシゾウムシ | ダンダラカッコウムシ | |||
| フィリピンキクイムシ | アリモドキカッコウムシ | |||
| マツノツノキクイムシ | ||||
| エタノールの影響なし | クロカミキリb | |||
| エタノール | ピネンの影響なし | コバケデオネスイ | シロヒゲナガゾウムシ | |
| サクキクイムシ | ||||
| ピネンが効果を弱める | キイロコキクイムシ | ヒメナガセスジホソカタムシ | ハンノキキクイムシ | |
| サクセスキクイムシ | ||||
| オノハラキクイムシ | ||||
11.マツノマダラカミキリの天敵サビマダラオオホソカタムシの枯死樹幹内における寄生状況
浦野忠久(昆虫研究室)
1. はじめに
サビマダラオオホソカタムシ(Dastarcus helophoroides Fairmaire、以下ホソカタムシ)はカミキリムシ類の捕食寄生者であり、野外におけるマツノマダラカミキリ(以下マダラ)への寄生は、1979年に広島県尾道市で初めて認められた。しかし岡山、広島県以外の地域からは、本種のマダラに対する寄生は今のところ報告されていない。過去におけるこれらの地域での寄生率は、19.5~58%であった。1999年、岡山県内のマツ林において、ホソカタムシの寄生したマツ枯死木を調査したので、野外における本種の寄生状況について報告する。
2. 材料と方法
調査地は岡山県和気郡和気町の36年生アカマツ林である。1999年5月中旬に林内の枯死木16本を伐倒し、樹幹を長さ1mに玉切りした。これらの丸太合計123本を5月21日に支所構内へ搬入し、野外の網室に入れて保存した。搬入した丸太を5月下旬から7月上旬にかけて割材し、材内の調査を行った。調査は丸太内(樹皮下、材内)のマダラ及びその天敵の種類、生息数及びステージについて行った。マダラについては、生存個体数のほかに材入孔数及び材内孔道数、成虫の脱出孔数を調べた。
3. 結果と考察
割材調査の結果を図に示した。マダラの天敵としてはホソカタムシ、オオコクヌスト及びキツツキ類が確認された。供試木内にはマダラの材入孔及び材内孔道が高密度に見られたが、それに比べて生存虫数は非常に少なかった。マダラの存在しない材内孔道の内で樹幹の繊維方向に孔道が伸長しているもの、および蛹室が形成されているものを、かつてそこに幼虫が存在したが捕食などの原因で死亡したものと仮定すると、そのような不在孔道が全体の78%を占めた。供試木からはオオコクヌストが合計19頭採集された。したがって不在孔道の一部は、本種の捕食によるものと考えられる。キツツキ類による捕食は樹幹表面の食痕から判別した。キツツキ類に捕食されたと見られる不在孔道は全体の7%であった。ホソカタムシはマダラの材内幼虫及び蛹に寄生していた。本種による寄生率は5月下旬にはわずか3%であったが、7月上旬には15%に達した。それとともに供試木内のマダラ生存個体数が減少し、5月下旬及び6月上旬には生存率10%以上であったものが、6月下旬には27本の供試丸太から、幼虫と蛹それぞれ1頭しか採集されなかった(生存率1%)。不在孔道とキツツキによる捕食を除いた寄生率(5月以降実際に材内に生存していたマダラの幼虫と蛹に対する寄生率)は、5月下旬の約20%から徐々に増加して最終的には80%前後となった。今回の調査ではホソカタムシの寄生率が調査期間を通じて上昇しており、網室内での幼虫の寄生時期は、マダラの羽化脱出時期とほぼ一致していた。したがってホソカタムシは、オオコクヌスト及びキツツキ類による捕食の後で、最終的な密度調節要因として働くことが明らかになったものと考えられる。
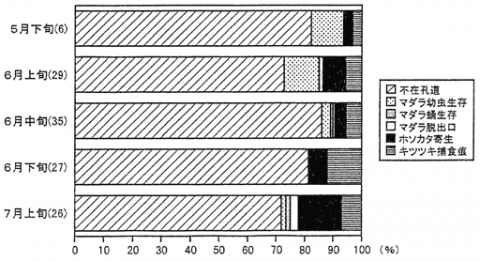
岡山県から採集した枯損マツにおける、マツノマダラカミキリ材内孔道の状況
(括弧内の数字はそれぞれの時期に割材調査した丸太の本数を示す)
12.シカとネズミがササの現存量及び樹木実生の生存におよぼす影響
日野輝明・島田卓哉(鳥獣研究室)
伊東宏樹(造林研究室)
古澤仁美(土壌研究室)
高畑義啓(樹病研究室)
上田明良(昆虫研究室)
1. はじめに
本研究の目的は、草食者のシカとネズミ、光や土壌養分をめぐる競争者のササの各要因が、実生の生存や成長に及ぼす直接的・間接的影響の動態を、操作実験に基づいて定量的に調べることである.本論では、実験開始後1年目と2年目の結果について報告する。
2. 方法
奈良県大台ヶ原の針広混交林内に、ネット等によるシカとネズミの除去の有無及びササの刈取りの有無の8通りの組み合わせからなる20mx20m実験区を5セット設置し、97年に調査を開始した.各処理区内に1m四方の樹木実生調査プロットを設置し、全個体について発生と生死の追跡調査を定期的に行った。ササは秋にシカとネズミの有無の組み合わせからなる4通りの各処理区において、6サンプルずつ(半径10cmの円)刈り取って持ち帰り、乾燥重量、桿密度、桿長、葉長、枝数、葉数を調べた。
3. 結果と考察
1年目、2年目ともに、シカの実験的除去が処理区内のササの桿長、葉長、枝あたりの葉数を増加させたのに対して、ネズミの除去はササの桿密度を増加させた(図-1)。処理区間で見られたこの除去効果の違いは、シカがササの枝葉部分を食害したのに対して、ネズミが新生桿を食害した結果だと推測できる.ササの現存量(乾燥重量)に対する除去の効果は、1年目にはネズミのみであったが、ササが伸長してきた2年目にはシカとネズミの両方に見られるようになった。今後はササの伸長にともなって、ササの現存量に及ぼすシカの除去の効果がネズミの除去の効果を上回っていくことが予想される。
広葉樹の実生の死亡要因として、1年目にはシカ、2年目にはササの影響が認められた。一方針葉樹の実生については、1年目にはどの要因の影響も見られなかったが、2年目にササとシカ両方の影響が認められた(図-2)。2年目になってササの効果が見られるようになったのは、ササの現存量の回復によるものと考えられる。このことは、ササの現存量増加にともなう実生の年間死亡率の増加が、広葉樹、針葉樹ともに2年目になって認められるようになったことからも裏付けられた(ケンドールの順位相関:広葉樹τ=0.478、針葉樹τ=0.462、p<<0.005)。ところが、シカとササあるいはネズミとササの両方の要因があるときの実生全体の死亡率は、ササ単独のときの死亡率よりも小さかった(U検定:シカp=0.081;ネズミp=0.011)。これはシカとネズミの採食によってササの現存量が減少したためであると考えられる(図-1)。つまり、シカとネズミは、直接的には採食により実生に対してマイナスの影響を与えるが、間接的には実生と競争関係にあるササを採食することでプラスの影響を与えていることが分かった。今後はササの現存量の回復にともなって、ササによる実生死亡率の増加、及びシカ・ネズミのササ採食による間接的な実生へのプラスの効果がさらに顕著になっていくことが予想される。
以上の結果から、樹木実生の生存・成長には、シカ、ネズミ、ササの直接的及び間接的な相互作用が関係しており、それはまたササの現存量に依存してダイナミックに変動するものであることが明らかになった。このようなデータの蓄積は、天然更新を促進し、かつ生物多様性の高い森林生態系を維持していくために、シカの密度管理やササの人為的な刈取りなどの施策を講じていく上で重要な資料となると考えられる。
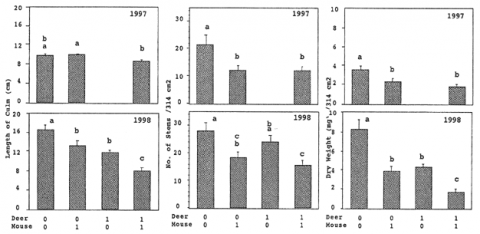
図-1 シカとネズミのあり(1)なし(0)がササの稈長、稈密度、乾燥重量の平均値(+標準偏差, N=30)に及ぼす影響. 図中の同じ記号は、Bonferroniの多重比較法により有意差がない(P>0.05)ことを示す.
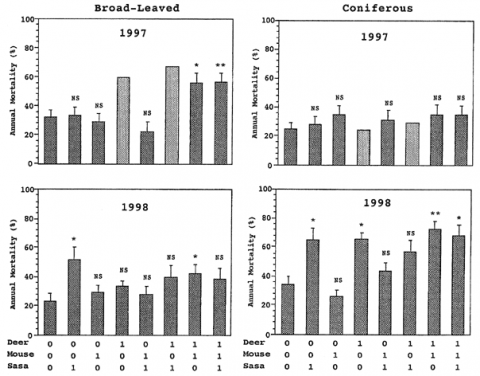
図-2 シカとネズミのあり(1)なし(0)が広葉樹と針葉樹の実生の年間平均死亡率(+標準偏差, N=10)に及ぼす影響. 97年の「100」「101」の値は、データが得られなかったため、シカとネズミの間に相互作用がない、効果は相加的に働くと仮定して求めた推定値. **P>0.01, *P<0.05, NS P>0.05: 「000」の値との差のU-検定.
Copyright © Forest Research and Management Organization. All rights reserved.